
Kromosommutation

Kromosommutationer er ændringer i dele af kromosomer, der omfatter mere end et gen. Kromosommutationer indebærer manglende eller tilføjede dele af kromosomalt DNA eller en uregelmæssig fordeling af dette. Det kan stamme fra et atypisk antal kromosomer eller en strukturel abnormalitet i en eller flere kromosomer. Kromosommutationer kan undersøges ved at sammenligne med en "normal" karyotype ved forskellige typer af genetiske undersøgelser. Baggrunden for kromosommutationer er som oftest fejl i celledeling i enten meiose eller mitose, og afhængigt af tidspunktet eller typen af denne mutation, kan den overføres til cellens afkom.
Typer[redigér | rediger kildetekst]
Der er mange typer kromosommutationer, og de inddeles efter deres kvalitative aspekter eller deres kvantitative aspekter.[1] Patogenesen af forskellige kromosommutationbetingede genetiske sygdomme afhænger af typen af mutationer, og om de vil kunne overføres til organismens afkom.[2]
Den kvalitative inddeling indordner kromosommutationerne efter den måde, hvorpå kromosomernes struktur er blevet forandret, mens den kvantitative inddeling inddeler disse mutationer efter om de ændrer på kromosommaterialets mængde.[2]
Numeriske mutationer[redigér | rediger kildetekst]

Numeriske mutationer er heteroploidier, dvs. hvor antallet af kromosomer ikke er det sædvanlige for den givne art (i mennesket er det 46).
Kopiering af hele karyotypen, der resulterer i en eksakt multiplicering af antallet af de haploide kromosomer (n), er en euploidi. I mennesket er antallet af haploide kromosomer 23 (n = 23). En euploidi, hvor antallet af haploide kromosomer er 3n, kaldes triploidi; 4n kaldes tetraploidi og så fremdeles. Triploidier ses i 1-3% af registrerede befrugtninger, hvoraf langt de fleste resulterer i en spontan abort, mens de få, der overlever fødslen, ikke overlever længe.[2] Velbeskrevne tilfælde af ikke-mosaiske, tetraploide børn, der overlever graviditeten, er ikke kendt.
Alle andre kromosomantal kaldes aneuploidier og forekommer i 5% af menneskelige graviditeter.[2] Af disse ses monosomier (hvor et kromosom kun har én haploid kopi). Et eksempel på en monosomi kompatibel med liv er monosomi-X (Turners syndrom), hvor karyotypen er 45,X. Desuden ses trisomier, hvoraf især X-kromosom-trisomier er kompatible med overlevelse af og efter fødslen. Trisomier i autosomale kromosomer har ofte mere alvorlige fænotyper, og især 3 er (i varierende grad) kompatible med overlevelse (i varierende perioder): trisomi-21 (Downs syndrom, karyotype 47,XX,+21 eller 47,XY,+21), trisomi-18 (Edwards syndrom, karyotype 47,XX,+18 eller 47,XY,+18) samt trisomi-13 (Pataus syndrom, karyotype 47,XX,+13 eller 47,XY,+13).[2]
Numeriske mutationers årsager er ikke fuldt kendt, men de fleste skyldes nondisjunction i enten 1. meiotiske deling, hvor bivalente ikke segregeres korrekt, hvilket medfører 1 kopi af hvert parentalt homologt kromosom, eller 2. meiotiske deling, hvor de to søsterkromatider ikke segregeres korrekt, hvilket resulterer i 2 kopier af samme parentale kromosom.[2]
Strukturelle mutationer[redigér | rediger kildetekst]

Strukturelle mutationer er omfordelinger af kromosomalmateriale, og de resulterer ikke i en ændring af antallet af kromosomer. De inddeles kvantitativt efter, om de ændrer på mængden af kromosommateriale eller ej.
- Balancerede strukturelle mutationer ændrer ikke på kromosommaterialets mængde
- Ubalancerede strukturelle mutationer ændrer på kromosommaterialets mængde.
Fælles for strukturelle mutationer er, at de ofte skyldes skæv genetisk rekombination (skæv overkrydsning) i meiosen.[2]
Deletioner[redigér | rediger kildetekst]
Deletioner er tab af kromosomalsegmenter, og de resulterer i tab af genetisk materiale. De er derfor altid ubalancerede strukturelle mutationer. Om de viser sig fænotypisk afhænger af den mængde genetiske materiale, der er blevet slettet, og hvad der blev slettet. Deletion af kodende segmenter af DNA kan resultere i haploidinsufficiens, hvor den tilbageværende mængde DNA fra det ikke-afficerede homologe kromosom ikke er tilstrækkelig til at opretholde fysiologisk funktion.[2] Dette ses f.eks. i Wolf-Hirschhorns syndrom (deletion af et stykke af den korte arm af kromosom 4, angivet som del(4p16.3) i cytogenetisk notation).[3]
Afhængigt af placeringen af deletionen på kromosomet kan de underinddeles i:[2]
- Terminale deletioner, hvor enden af et kromosom slettes.
- Interstitielle deletioner, hvor en intern del af kromosomet slettes.
Duplikationer[redigér | rediger kildetekst]
Duplikationer involverer kopiering af kromosomalsegmenter, der resulterer i ekstra genetisk materiale. De er derfor også altid balancerede strukturelle mutationer. Om de viser sig fænotypisk afhænger af den mængde genetiske materiale, der er blevet duplikeret, og hvad der blev duplikeret. Duplikation inde i et kodende segment, f.eks. et gen, kan eksempelvis forstyrre dennes genekspression og resultere i en patologisk fænotype.
Translokationer[redigér | rediger kildetekst]

Translokationer involverer en overflytning af kromosomalsegmenter mellem to forskellige kromosomer. Disse kan være meget forskellige i deres kvalitative natur, og de underinddeles derfor i reciprokke og robertsonske translokationer.
Translokationer er balancerede strukturelle mutationer, men hvis disse translokationer sker i kønscellelinjen kan en ubalanceret translokation nedarves ved, at kønscellens kromosomer ikke kan opstilles korrekt i meiosens metafase, hvilket ses på tegningen til højre, hvorfor de ofte segregeres forkert i meiosens anafase og får ikke genskabt deres oprindelige struktur. I stedet denne forkerte segregation medføre, at gameten får et derivativt kromosom, der indeholder segmenter af et andet kromosom, men ikke det segment, det burde have fået. Dette sker i 1-20% af tilfælde af de balancerede translokationer i kønscellelinjen.[2]
I somatiske celler er translokationer ofte ikke forbundet med sygdom, medmindre de flytter kromosommateriale på en måde, så en kodende sekvens af DNA brydes. Det anslås, at 1 ud af 500 mennesker har uopdagede translokationer.[2]
Reciprokke translokationer[redigér | rediger kildetekst]
Denne type sker ved brud på og rekombination af non-homologe kromosomer. Et stykke DNA fra et kromosom bytter plads med et stykke DNA fra et andet kromosom.
Robertsonske translokationer[redigér | rediger kildetekst]
Disse translokationer er de hyppigste translokationer og sker mellem to akrocentriske kromosomer (dvs. kromosomer, hvis centromer er tæt på kromosomets ender – i mennesket er det kromosom 13, 14, 15, 21, 22). De akrocentriske kromosomers p-arm (korte arm) indeholder mest satellit-DNA og hundredvis af kopier af rRNA-gener. Da disse forekommer i så mange kopier på alle de akrocentriske kromosomer vil deletioner af disse ikke resultere i en patologisk fænotype. I robertsonske translokationer vil to akrocentriske kromosomer smide deres p-arm, og deres respektive q-arme (lange arme) vil fusionere med hinanden og danne et derivativt kromosom. Der kan enten være én centromer i dette kromosom eller sjældnere 2. De smidte p-arme tabes.[2]
Insertioner[redigér | rediger kildetekst]
Insertioner er balancerede strukturelle mutationer. De kan enten betragtes som translokationer, fordi et kromosomsegment fjernes fra et kromosom og indsættes i et andet eller som en kategori af strukturelle mutationer for sig, fordi de ikke indebærer, at de bytter plads med et andet kromosomsegment. De er sjældne og resulterer kun i en abnorm fænotype, hvis kromosombruddene sker i et kodende kromosomsegment. Hvis denne mutation sker i kønscellelinjen er der dog risiko for, at gameterne arver en ekstra kopi af et kromosomsegment eller en kopi for lidt. I mennesket vil det hhv. betyde en funktionel trisomi og funktionel disomi for det afficerede segment.[2]
Inversioner[redigér | rediger kildetekst]
Disse mutationer sker, når der sker 2 dobbelstrengsbrud i samme kromosom, og disse brud ved nonhomolog rekombination bliver rekombineret, således at segmentet rekombineres omvendt af, hvad det var oprindeligt.
- Pericentriske inversioner sker, hvis det ene dobbeltstrengsbrud er sket i p-armen og det andet er sket i q-armen. Herved bliver centromeret også vendt om.
- Paracentriske inversioner sker, hvis begge dobbeltstrengsbruddene er sket i den samme kromosomarm. Segmentet vendes om, men centromeret vendes ikke om.
For begge disse inversioner gælder det, at de ofte ikke fører til abnorme fænotyper, men hvis de sker i kønscellelinjen kan abnorm overkrydsning ske. Både de pericentriske og paracentriske inversioner kan ved meiosen danne gameter med kromosomer med ubalancerede segmenter, men kun de paracentriske inversioner kan medføre, at de ubalancerede kromosomer i gameterne indeholder 2 (dicentriske kromosomer) eller ingen centromerer (acentriske kromosomer). Disse er ofte inkompatible med liv.[2]
Ringkromosomer[redigér | rediger kildetekst]
Disse mutationer forekommer i ca. 1 ud af 2500 mennesker og er karakteriseret ved, at et kromosom i stedet for at have den normale lineære form (i eukaryoter) bliver forbundet med sin ende og danner et ringformet kromosom (ringkromosomer er dog normale i prokaryoter). Dette kan skyldes tab af telomerer, der fører til en rekombination af den ene kromosomarm med den modsatte kromosomarm. Sådanne kromosomer kan forstyrre celledeling, og afhængigt af deres egenskaber kan de medføre varierende grader af forstyrrelser af fænotypen.[2]
Isokromosomer[redigér | rediger kildetekst]
Isokromosomer er kromosomer, hvis ene arm er blevet tabt, og den resterende arm er blevet kopieret og indsat på den tabte arms plads som et spejlbillede. Dette resulterer i, at der kommer en ekstra kopi af det genetiske materiale på den kopierede arm (i mennesket funktionel trisomi), mens der kommer en kopi for lidt af den tabte arm (i mennesket funktionel monosomi). Sådanne isokromosomer ses ofte i neoplastiske celler (herunder kræftsvulster og visse former for blodkræft).[2]
Nedarvning hos mennesker[redigér | rediger kildetekst]
Kromosommutationer kan være nedarvet fra en forælder eller ske de novo, dvs. i den organisme, hvor kromosommutationen optræder. De novo-kromosommutationer kan ske både i den somatiske cellelinje og i kønscellelinjen.
Eftersom den somatiske cellelinje og kønscellelinje adskilles fra hinanden i den embryonale udvikling, vil en de novo-mutation i somatiske celler ikke blive nedarvet til personens afkom. Til gengæld kan den, hvis dens egenskaber taler herfor, vise sig ved en abnorm fænotype hos patienten med mutationen.
Såfremt de novo-mutationen sker i kønscellelinjen, er der risiko for, at anomalien overføres til gameterne (kønscellerne) og nedarves til personens afkom, ofte resulterende i spontan abort eller nedsat fertilitet. Til gengæld kan en mutation i kønscellelinjen ofte ikke medføre mærkbart abnorme fænotyper hos personen med mutationen, idet den somatiske cellelinje ikke er påvirket.
En de novo-mutation i mitosen i embryonale stamceller tidligt i fosterudviklingen kan medføre mosaicisme, dvs. en tilstand, hvor en person har forskellige celler med forskellige karyotyper. Jo senere denne mutation sker i fosterudviklingen, des færre celletyper vil kromosommutationen påvirke. På denne måde kan kromosomalt betingede genetiske sygdomme vise sig med varierende fænotyper. Eksempelvis kan personer, der er mosaikker for trisomi-21 have en signifikant højere IK end personer, hvis alle celler har trisomi-21.[4] Mosaicisme i kønsceller (kønscellemosaicisme), hvor nogle kønsceller har en mutation, mens andre ikke har, kan medføre, at en person med en mutation giver mutationen videre til nogle børn, men ikke andre børn. Dette kan ses i en abnormt stor rate af spontane aborter, og ofte udføres genetisk rådgivning derfor hos par, der ikke kan opnå en vellykket graviditet, for at undersøge, om en kromosommutation hos en eller begge patienter er årsagen.
Erhvervede kromosommutationer[redigér | rediger kildetekst]
De fleste cancertyper, hvis ikke alle,[5] forårsager kromosommutationer, hvor gener forstyrres. Ofte er årsagen til cancers invasive egenskaber mutationer, der fører til overekspression af protoonkogener eller nedregulering af tumorsupressorer. Dette kan eksempelvis ske ved fusion af gener, således at et normalt undertrykt protoonkogen reguleres af en anden, mere udtrykt regulator, eller et tumorsupressor-gen, der fusioneres med et gen, der styres af en mindre undertrykt regulator.[6]
Skitser over nogle kromosommutationer[redigér | rediger kildetekst]
-
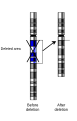 Skitse af en deletion, hvor et kromosomområde tabes.
Skitse af en deletion, hvor et kromosomområde tabes. -
 Genduplikation indebærer, at der kommer flere kopier af et kromosomsegment.
Genduplikation indebærer, at der kommer flere kopier af et kromosomsegment. -
 Reciprok translokation.
Reciprok translokation. -
 Insertion.
Insertion. -
 Paracentrisk inversion. En pericentrisk inversion ville have omfattet centromeret, hvilket ikke er tilfældet her.
Paracentrisk inversion. En pericentrisk inversion ville have omfattet centromeret, hvilket ikke er tilfældet her. -
 Formation af isokromosomer.
Formation af isokromosomer.





Se også[redigér | rediger kildetekst]
Referencer[redigér | rediger kildetekst]
- ^ Atlas of Genetics and Cytogenetics in Oncology and Haematology: Chromosomes, Chromosome Anomalies. Tilgået 31.12.2016 på: http://atlasgeneticsoncology.org/Educ/PolyMecaEng.html
- ^ a b c d e f g h i j k l m n o p Nussbaum, Robert L.; McInnes, Roderick R.; Willard, Huntington (2016). Thompson & Thompson Genetics in Medicine (8 udgave). Philadelphia, Pennsylvania: Elsevier. s. 64-85.
{{cite book}}:|access-date=kræver at|url=også er angivet (hjælp) - ^ Dufke, A., Seidel, J., Schöning, M., Döbler-Neumann, M., Kelbova, C., Liehr, T., . . . Enders, H. (2001). Microdeletion 4p16.3 in three unrelated patients with Wolf-Hirschhorn syndrome. Cytogenetic and Genome Research, 91(1-4), 81-84. doi:10.1159/000056823. Tilgået 31.12.2016 på: https://www.ncbi.nlm.nih.gov/pubmed/11173835
- ^ Batshaw, Mark, red. (2005). Children with disabilities (5th udgave). Baltimore [u.a.]: Paul H. Brookes. s. 308. ISBN 978-1-55766-581-2.
- ^ Atlas of Genetics and Cytogenetics in Oncology and Haematology. Tilgået 31.12.2016 på: http://atlasgeneticsoncology.org/Educ/Hempat_e.html
- ^ "Mitelman Database of Chromosome Aberrations and Gene Fusions in Cancer". Arkiveret fra originalen 25. maj 2016. Hentet 31. december 2016.